|
|
||||||
| by M.A.Fusco | ||||||
|
|
||||||
|
The term proprioceptive sensitivity was introduced for the first time in 1906 by Sir Sherrington to indicate the set of nervous signals originating within the organism during the movement. According to this definition suitable stimuli are conveyed from the organism to specific receptors located within the organism that's why they are called proprioreceptors (that means own receptors). Almost 100 years after this definition was coined, it is still sound, even if the concept of sensitive proprioceptivity gained new specific meanings. At the beginning proprioceptive sensitivity concerned only signals coming from the neuromuscular spindles and from the musculo-tendineous corpuscles of Golgi. Nowadays it also concerns stimuli coming from tactile, pressure, sore receptors etc.. The main and most studied proprioceptors are the neuromuscular spindles and the Golgi's musculo-tendineous corpuscles. Both receptors belong to the class of stretching receptors because they are sensitive to physical variations caused by the stretching of the fibres of the muscle. |
||||||
|
|
||||||
|
STRUCTURE AND FUNCTION OF THE NEUROMUSCULAR SPINDLES |
||||||
|
|
||||||
|
||||||
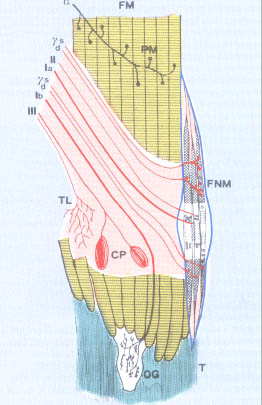
| The modified muscles fibers that build up the spindle are called intrafusal fibers and are thinner than the real muscle fibers. Normally intrafusal fibers are classified into two types: | ||||||
|
||||||
The nuclear bag fibers are furthermore classified into two
subtypes:
|
||||||
|
Their structure, their biochemical characteristics and their
mechanical features are quite different. The innervation of the
spindle is very complex. Nerve fibers of the spindle are both sensory and motory. Sensitive afferent fibers are divided into:
|
||||||
|
Primary afferent fibers have a high speed of conduction and measure from 12
up to 20 micromm in diameter. These fibers belong to the first group of the
classification of the nervous fibers according to Llyod. They originate
prevalently from spiral endings twisted around the central part of the
intrafusal fibers of the three above described types. Secondary afferent fibers have a lower speed of conduction and a shorter diameter. They belong to the second group of the classification of the nervous fibers according to Llyod. They originate prevalently from spiral endings twisted around the central part of nuclear chain fibers. There are also the so-called flower endings that are located almost exclusively within the nuclear bag fibers. A particular characteristic of the fusal receptor is represented by the motory fibers coming from the gamma motorneurons of the ventral horn of the spinal cord. These fibers are classified into two types: |
||||||
|
1. Dynamic gamma fibers, 1 or 2 on each side, innervate the polar region of the dynamic nuclear bag fibers; 2. Static gamma fibers, they innervate both static nuclear bag fibers and nuclear chain fibers. |
||||||
|
|
||||||
| In order to understand how the spindle can inform about the variations of muscle length, we must above all focus on the anatomic and histological position of the spindles within the muscles tissue. The capsule of the spindle melts, at the level of its two poles, with the connective sheath that surrounds the extrafusal fibers. Thanks to this parallel orientation each lengthening of the muscle is transmitted also to the intrafusal fibers, in the same way, a stretching of the muscle causes a spacing of the spirals of the nervous ending twisted on the central part of the intrafusal fibers where there is a gelatinous fluid that that probably exerts a lubrificating action and allows some independent movements among the fibers. We have also to focus on the fact that neuromuscular spindles are receptors of first type. That means that they are able to react to the variations of incidental energy through an electric potential called Generator Potential (GP). The stimulus produces an electronic variation of the potential of the membrane that is quite similar to the exciting post-synaptic potential. It directly depends from the width of the stimulated area and from the intensity of the stimulus and it is caused by a non-selective modification of the permeability of the membrane to all ions. Above all ions Na+ pass through this membrane because electronic and chemical forces push them, in this way they create an intracellurar flux of depolarisation positive charges. The deformation triggers the process of mechanoelectric transition and the depolarisation of the nervous ending by transmitting the potential of action to the first knot of Ranvier of the afferent fiber. The neuromuscular spindle is highly sensitive to the stretching stimulus, very short variations of the muscle length cause a big variation of the frequency of the discharge of the afferent fibers. Thanks to the presence of two different types of intrafusal fibers the nervous system gets information both on the size of the lengthening of the muscle through the variations of the discharge of the secondary fibers and on the speed of the lengthening of the muscle through the fibers of group Ia. During the contraction of extrafusal fibers, neuromuscular spindles, due to their parallel arrangement, undergo a shortening that inhibits and stops the afferent discharge. The control over the sensitivity of the receptor is exerted by the alfa and gamma innervation system. The gamma innervation produces a contraction of the two poles of the intrafusal fibers that on its turn causes a stretching of the central part of these fibers; the nervous endings are twisted around this part. In this way the detention of the spindle produced by the contraction of the extrafusal fibers caused by alfa motoneurons can be counterbalanced by the opposite effects produced by the activation of the gamma motoneurons that occurs at the same time. So the sensitivity of the receptors can be constant at each level of muscle contraction, furthermore the control of the sensitivity of the receptor carried on by the gamma system can selectively increase the static and dynamic sensitivity of the spindle through the two types of gamma fibers that innervate it. Impulses coming from the spindles reach the spinal cord and excite the alfa motoneurons and the motoneurons of the antagonist muscles through a monosynaptic transmission. This is the afferent branch of the myotatic reflex that means that the active and passive stretching of the muscle triggers, by activating the fusal proprioreceptors, a reflex contraction that is able to restore the initial length of the muscle. Myotatic reflex occurs both in extensor and in flexor muscles, and above all in the anti-gravitational muscles that play the most important role in maintaining the posture. In these cases the stimulated component of the spindle is the static one. There are higher spinal centres of control of the tonic gamma motoneurons that are located in the reticular structure and within the set of the vestibular nuclei. These centres optimise the functional importance of the proprioceptive system of the neuromuscular spindles. The secondary afferences, fibers belonging to group II, accomplish a different task. They come from the ring-spiral shaped endings located on the intrafusal nuclear chain fibers. They carry out a monosynaptic function and exert very complex exciting polisynaptic effects on flexor muscles and inhibiting effects on extensor muscles. So fibers belonging to group II contribute in building up the tonic component of the myotatic reflex, because they are sensitive to the static muscle stretching; they excite the flexor component of the spinal motoneurons through polysinaptic connections; under a condition of hyper tonus, such as spasticity, their inhibiting stimulation increases as consequence of the passive stretching exerted by the muscle. | ||||||
|
|
||||||
|
STRUCTURE AND FUNCTION OF THE MUSCULO-TENDON ORGANS OF GOLGI |
||||||
|
|
||||||
|
||||||
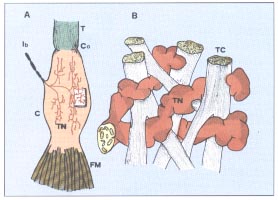
| Muscular fibers related to the organ of Golgi belong to different motory units and this allows the receptors to test and select the tension developed by different kinds of motory units during the contraction of the muscle. Unmyelinated irregular shaped nervous endings penetrate into connective trabecules of the tendon. The development of tension of the muscle stretches the filaments that become straighter. This mechanical deformation depolarises the filaments firing the action potential in the extracapsular myelinated part of the nervous fiber. Impulses reach the spinal cord passing through high-speed conduction fibers of group Ib. They exert an inhibiting action on the alfa motoneurons and on the antagonist muscles through just one spinal interneuron. These organs are extremely sensitive to the active contraction of the muscles and they are almost indifferent to the passive stretching of the same muscles. In the past the organs of Golgi were considered as a defence of the organism able to detect and inform about excessive contraction and prevent crash of the muscular insertions. Nowadays, on the contrary, we know that a massive activation of these afferences can stop the muscular contraction, but their main function is to inform the higher centres about the average tension developed by a selected group of motory units. The discharge frequency increases with the progressive increasing of the amount of the involved motory units. This is a mechanism of negative of feedback control for the contraction. The organism uses this mechanism, in order to allow the motory centres to program the contraction force. | ||||||
|
|
||||||
|
UPPER SPINAL PROJECTIONS OF MUSCULAR PROPRIOCEPTORS |
||||||
|
|
||||||
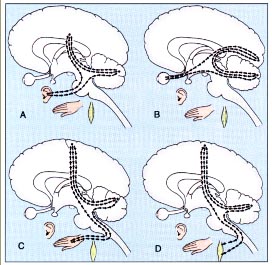
| Muscular proprioceptors convey information to different upper spinal structures involved in the programmation and in the control both of statics and of movement. The most informed structures are the cerebellum and the sensitive-motory cerebral cortex. | ||||||
|
The main afferent pathways to the cerebellum are four: 1. Dorsal spinocerebellar tract 2. Ventral spinocerebellar tract 3. Cuneocerebellar tract 4. Rostrocerebellar tract |
||||||
|
||||||
| Fibers of the spinocerebellar tract terminate in the anterior ipsilateral cerebellum as mossy fibers.The ventral spinocerebellar tract conveys in the same region of the anterior ipsilateral cerebellum information coming above all from the muscolo-tendineous organs of Golgi and is a monitoring and control system of the muscular tension developed in the muscles of the lower limbs. | ||||||
|
||||||
| corresponding to the area 3a of Brodmann; -proprioceptive afferences coming from the lower limbs through the lateral cordons of the spinal cord, the Z nucleus of the encephalic trunk and of the thalamus. | ||||||
|
|